La transición del estado preñada no lactante al no preñado lactante es un período de cambios drásticos para la vaca, la cual debe adaptar su metabolismo a las fuertes exigencias que le demanda la producción. Del equilibrio con que la vaca resuelva este proceso dependerá la capacidad de maximizar la producción y la calidad de la leche, de evitar enfermedades metabó- licas y asegurar la siguiente preñez. La mejora nutricional, la selección genética y el manejo animal han aumentado la producción de leche en las últimas décadas, y esto se asocia a una disminución del desempeño reproductivo y al aumento de problemas sanitarios. Esta revisión resume trabajos realizados en los últimos años en Uruguay, enfatizando en el manejo diferencial de nutrientes, la respuesta en comportamiento ingestivo, los mecanismos endocrino-moleculares de la partición de nutrientes y su relación con la fertilidad en vacas lecheras. Se reportan experimentos que estudian la eficiencia productiva de diferentes biotipos lecheros y razas. Se describen trabajos en salud que identifican el período de transición como período de riesgo a enfermedades metabólicas, infecto-contagiosas y traumáticas. Se concluye que los estudios integrados en problemas de relevancia nacional es la respuesta necesaria a sistemas biológicos complejos como lo es la vaca lechera durante el período de transición en pastoreo.
Producción de leche en Uruguay
En Uruguay, la producción de leche se ha incrementado en los últimos 30 años; sin embargo, el número de productores ha descendido significativamente, despareciendo más de 2000 productores en los últimos 20 años, concomitantemente con un aumento de la escala de los tambos en términos de número de vacas en ordeñe (DIEA, 2011). Las exportaciones de productos lácteos se aproximan al 70% en relación al volumen total disponible, fundamentalmente leche en polvo y quesos, por lo que cobra relevancia el contenido de sólidos de la leche. Consecuentemente las industrias han promovido el criterio de pago por sólidos. Estas condiciones son inmejorables para sentar las bases y generar las herramientas necesarias que permitan agregar valor a la materia prima para la elaboración de productos básicos especializados para nuestros mercados compradores.
En nuestro país, por tratarse de un sistema pecuario de base pastoril, la producción de leche acompaña la producción estacional de las pasturas, dándose en primavera el máximo de producción de leche (DIEA, 2011). Los sistemas de producción de leche exhiben un desbalance estructural entre oferta y demanda de nutrientes. Este desbalance se intenta corregir con suplementación, con reservas forrajeras y concentrados, derivando en sistemas con niveles crecientes de complejidad operativa, requerimientos de infraestructura y fundamentalmente de precisión en el manejo de los recursos alimenticios. El uso intensivo de pasturas mejoradas con concentrados y ensilados fueron uno de los factores clave que permitieron un salto productivo en las dos últimas décadas (DIEA, 2011). Las evaluaciones genéticas nacionales de la raza Holando indican que la genética del rodeo lechero uruguayo proviene predominantemente de América del Norte. La selección genética por producción de leche asociada a la difusión del Holando de alta producción- a nivel mundial durante las últimas décadas ha sido un factor primordial en la mejora productiva internacional.
Aspectos limitantes del sistema de producción lechero uruguayo
La leche bovina juega un papel central en la nutrición humana y es un ingrediente que se consume globalmente en diferentes formas y destinos. Las principales líneas de investigación de la industria láctea internacional se focalizan en la valorización e innovación de productos por las propiedades de algunos componentes lácteos promoviendo los aspectos saludables y terapéuticos de la leche (Bauman et al., 2006). Por ejemplo, el ácido linoleico conjugado (CLA) tiene propiedades como inhibidor de la carcinogénesis y de la aterogénesis, mejora la capacidad del sistema inmune, previene la obesidad y posee efectos antidiabéticos y mejoras en la mineralización ósea (Pariza et al., 2001). Existen numerosos reportes al respecto, sin embargo, no se ha llegado a una conclusión final respecto de la seguridad y beneficios del CLA (Benjamín y Spener, 2009). El aumento en la ingesta de la pastura fresca resulta en un aumento de 2 a 3 veces en el contenido CLA en la leche (Dhiman et al., 1999) y esto posiciona a nuestro sistema pastoril de producción de leche de forma ventajosa a nivel internacional. Actualmente el agregado de alguno de estos elementos en la leche se realiza a nivel industrial. La investigación nacional en calidad de leche en Uruguay respecto a la maximización de componentes nutracéuticos de forma natural ha sido escasa. Resulta entonces prioritario que Uruguay como «Marca País» genere un mensaje claro, sólido, inteligente y coherente acorde a esa imagen.
Por otro lado, el consumo de materia seca (MS) en sistemas de pastoreo es usualmente más bajo que en sistemas de confinamiento y podría ser insuficiente para sostener la alta producción de leche que podría lograrse con el potencial genético (Kolver y Müller, 1998; Chilibroste et al., 2012a). En investigaciones nacionales el análisis de las curvas de lactancia sugiere que los animales no logran expresar su potencial productivo, seguramente en respuesta al desacople entre requerimientos-oferta de nutrientes y ambiente productivo (Chilibroste et al., 2012a). A la complejidad mencionada del sistema pastoril de producción de leche se le debe agregar la escasa comprensión del comportamiento ingestivo de la vaca lechera durante la transición. Esto es especialmente relevante debido a la depresión del consumo que ocurre en las últimas semanas previas al parto (Grummer, 1995).
Producir un ternero a intervalos regulares es un prerrequisito para obtener una performance rentable de la lactación (Royal et al., 2002) y el intervalo parto concepción (IPC) es el factor limitante. El aumento productivo ha provocado una disminución de los indicadores reproductivos; la información es consistente en señalar una disminución de 20 a 30% en las tasas de preñez desde la década del ´60 al presente en diferentes países y un aumento de problemas reproductivos y sanitarios (Roche et al., 2000; Lucy, 2001; Royal et al., 2002). En Uruguay, estudios en 200 mil lactancias indicaron que el IPC aumentó de 131 a 150 días de 1997-2001 a 2001-2005 (Rovere et al., 2007). Esto indica que la situación nacional actual no escapa a la de Holstein a nivel mundial, donde la presión por selección para producción de leche ha afectado los índices reproductivos. El síndrome de subfertilidad surge de la interacción del sistema de manejo, el biotipo y los procesos metabólicos que subyacen en ese aumento de la producción (Gutierrez et al., 2006). Como forma de levantar estas restricciones, en los últimos años se ha introducido genética de Nueva Zelanda, debido a la similitud del sistema pastoril con el de nuestro país. Nueva Zelanda exporta alrededor del 90% de la producción, por lo que cobran importancia los sólidos en leche. La producción en Nueva Zelanda prioriza el manejo de pasturas -90% de la dieta- por lo que la mayoría de los predios presentan parición estacional, y se procuran vacas livianas, sanas y fértiles. A nivel nacional, la investigación al respecto es incipiente. Hay aún menos información con respecto a la asociación de marcadores moleculares genéticos (por ejemplo: cambios de un solo nucleótido) y parámetros productivos y reproductivos en leche, a pesar de que estos kits comerciales están en Uruguay y se utilizan para predecir comportamientos fenotípicos de interés.
El conflicto en el desempeño productivo vs el reproductivo está en gran parte condicionado por el manejo diferencial en el que se puede incidir especialmente durante el período de transición. A la alta demanda metabólica por producción de leche se le suma la disminución (~ 30%) del consumo previo al parto (Grummer, 1995) que promueve la movilización de reservas corporales, es decir, el balance energético negativo (BEN). Los cambios que ocurren durante este período están directamente relacionados a los procesos de adaptación del sistema digestivo a la alimentación que recibirán luego del parto y del metabolismo. Los cambios en el metabolismo de los tejidos/órganos del cuerpo necesarios para apoyar una función fisiológica específica (homeorhesis, Bauman y Currie, 1980), aseguran la uniformidad del flujo de nutrientes en apoyo de la lactancia. Esta partición de nutrientes es comandada por señales hormonales que en conjunto con los perfiles metabólicos pueden ser utilizados como herramientas predictivas del estado de salud de rodeo. En ese estado fisiológico las vacas lecheras de alta producción tienen una utilización de nutrientes por parte de la glándula mamaria superior a la del resto del cuerpo, a tal extremo que Brown (1969) ha sugerido que la vaca debe ser concebida como un apéndice de la ubre y no viceversa. Este tiempo de sub-alimentación experimentado por los animales al inicio de la lactancia no solo afecta la magnitud de la respuesta residual de producción en toda la lactancia, sino que agrava el desempeño reproductivo. Es en este período que se concentran las enfermedades metabólicas o tecnopatías de la producción de leche actual. Del equilibrio con que la vaca resuelva este proceso dependerá la capacidad de maximizar la producción de leche, evitar enfermedades metabólicas y asegurar la siguiente preñez (Grummer, 1995).
La fragilidad del organismo se refleja en la actividad del sistema inmune de la vaca que está fuertemente deprimida alrededor del parto. Se ha descrito una disminución transitoria importante de células de defensa después del parto causado por un importante pasaje de neutrófilos hacia el tracto reproductivo. La capacidad de los linfocitos para responder y la producción de anticuerpos se ve también afectada alrededor del parto (Kehrli et al., 1989), por lo que se reportó una disminución dramática de los niveles séricos totales de inmunoglobulinas G y M en el período entre la semana 8 previa al parto y la cuarta semana postparto (Herr et al., 2011). Aún se desconoce el mecanismo exacto que determina la depresión del sistema inmunológico en el periparto pero se acepta que factores endócrinos y nutricionales estarían fuertemente involucrados (Goff y Horst, 1997; Vangroenweghe et al., 2005). Estos hallazgos podrían explicar la alta incidencia de enfermedades infecciosas durante este período.
Finalmente, la presión por producción y el manejo a gran escala de los tambos ha aumentado significativamente los factores de riesgo que desencadenan varias de estas enfermedades. En esas circunstancias, el período de adaptación de las vacas a la dinámica de ordeñe y cambios de manejo de la alta producción se puede ver comprometido. Pautas de comportamiento animal y diseño de instalaciones comienzan a jugar un importante rol como factores de riesgo de enfermedad y daño del bienestar animal. Uno de los problemas es el mantenimiento de los caminos y áreas de pasaje, así como los largos períodos que las vacas permanecen sobre el cemento de la sala de espera y las largas distancias que recorren para comer; esto representa un gran potencial traumático para el aparato locomotor. Por otra parte, la alimentación de estas máquinas metabó- licas para producir grandes volúmenes de leche implica un gran desafío nutricional. En nuestras condiciones, las grandes variaciones en la disponibilidad de diferentes tipos de granos y la inestabilidad de los precios conspiran contra una adecuada planificación de la nutrición de las vacas y exponen al sistema a riesgos sanitarios. Cambios bruscos y desajustes en estos aspectos pueden ser el origen de brotes de acidosis, cojeras y otros tipos de patologías que causan gran dolor, daño al animal y merma en la producción del rodeo.
Es por lo tanto el período de transición donde se concentran los problemas sanitarios del rodeo lechero y la base radica en la presión metabólica que implica los altos niveles de producción de leche potenciales del rodeo lechero actual desfasado de la capacidad de ingesta requerida. La clínica individual carece de efecto preventivo y representa un enfoque erróneo de las problemáticas sanitarias. Cuando las vacas se enferman, la leche ya se perdió y la intervención clínica –si bien necesaria– supone un esfuerzo tardío que en ocasiones tiene un magro impacto en controlar las pérdidas económicas que representan estas patologías (Ramos, 2007). Además, la buena imagen de la industria láctea pastoril asociada al bienestar animal y productos saludables se ve comprometida por la presencia de enfermedades en los rodeos lecheros actuales. El desafío de los sistemas de producción de leche al que se exponen los técnicos asesores no es posible de ser alcanzado por una sola disciplina. La necesidad de conocimiento original que permita comprender los procesos biológicos que subyacen a la relación ambiente-animal-producto es esencial para maximizar la relación costo-beneficio de la empresa. Esto es aún más relevante si se tiene en cuenta que la investigación de estos procesos se concentran en sistemas de estabulación y que el sistema pastoril de producción de leche sudamericano tiene características particulares que limitan la transferencia del conocimiento generado en otros sistemas productivos, incluso pastoriles.
Modo conceptual de trabajo
En la Figura 1 se esquematiza la forma de trabajo propuesta para generar conocimiento original en lechería. Todos los experimentos han sido realizados en vacas lecheras de raza Holando, con la excepción de los experimentos en donde hemos abordado el estudio de líneas genéticas y cruzas donde se especifica qué biotipos lecheros se han estudiado. Los recuadros indican los efectos o tratamientos (inputs) estudiados; el énfasis se ha puesto en manejo nutricional antes o después del parto, referido tanto a niveles de suplementación u oferta de pastura como tipo de nutriente. Hemos estudiado aspectos del manejo animal (hora de pastoreo, dominancia) y caracterizado las respuestas acorde a la categoría animal (edad, número de partos), e iniciado estudios respecto a la genética animal (cruza y biotipo lechero). Las variables de respuesta (outputs) económicas implican el producto (leche: cantidad y calidad en términos de grasa/proteína y de fracciones de caseínas y ácidos grasos) o parámetros reproductivos que son más difíciles de cuantificar (longitud del anestro posparto, tasas de preñez).
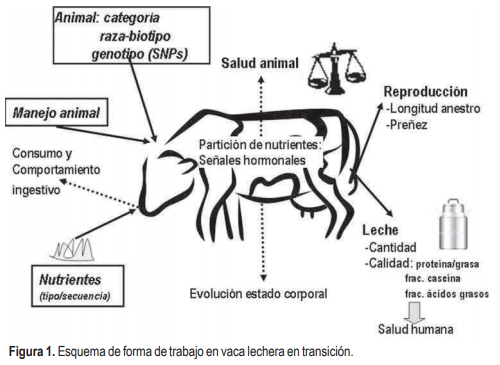
Las variables de respuesta que implican determinaciones en el animal como lo son el comportamiento ingestivo, la evolución de las reservas corporales (condición corporal, CC), los perfiles hormonales y metabólicos en sangre y la sensibilidad y/o respuesta de tejidos específicos permiten comprender cómo los diferentes inputs (efectos que se desean investigar) provocan los outputs. El estudio integrado de los mecanismos que implican la respuesta productiva a manejos animales diferenciales constituye el camino al desarrollo de tecnologías a ser transferidas a nuestro medio productivo. La investigación debe contemplar la repercusión a diferentes niveles de respuesta para así maximizar la rentabilidad del sistema productivo lechero, teniendo como eje primordial el mantenimiento de la salud y bienestar de los animales.
Período de transición: datos nacionales
En los experimentos realizados hemos observado que los perfiles metabólicos y endocrinos evidencian los cambios en CC durante el período de transición (Figura 2). La gran movilización en CC que ocurre en el pre y posparto temprano se acompaña de una pronunciada elevación de ácidos grasos no esterificados (NEFA) (Meikle et al., 2004; Cavestany et al., 2005, 2009a; Adrien et al., 2012; Ruprechter et al., 2011), que se asociaron con una mayor abundancia de los transcriptos en hígado de enzimas reguladoras de la β-oxidación (Carriquiry et al., 2010). A este aumento de NEFA le sigue frecuentemente un aumento de b-hidroxibutirato (BHB) el cual refleja el déficit energético (Figura 2A); si bien hay que recordar que pueden existir niveles elevados de BHB de origen ruminal debido a la hidroxilación del butirato ruminal.
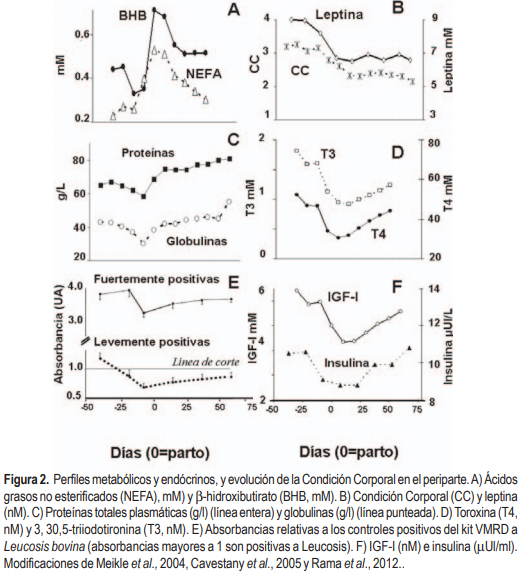
La pérdida de CC es en general más abrupta en vacas primíparas que multíparas (Meikle et al., 2004, 2005, 2006; Adrien et al., 2012) y se acompaña con una disminución en las concentraciones de leptina (Figura 2B), lo cual es consistente ya que esta es sintetizada por los adipocitos y varía con cambios en el porcentaje de depósitos grasos (Delavaud et al., 2000). Las vacas lecheras frecuentemente pierden más del 60% de su grasa corporal durante la lactación temprana (Tamminga et al., 1997). La leptina inhibe el consumo y aumenta la tasa metabólica, por lo que se sugiere que esta disminución es estratégica para estimular el consumo de la vaca lechera posparto y para disminuir el consumo periférico de nutrientes (disminución de la tasa metabólica). En el mismo sentido las hormonas tiroideas (T3, T4) disminuyen drásticamente antes del parto y las concentraciones observadas durante el posparto no se recuperan comparativamente con las del preparto (Meikle et al., 2004).
Los metabolitos nitrogenados disminuyen alrededor del parto y están vinculados a la dieta (Cavestany et al., 2005, 2009a; Meikle et al., 2004, 2006; Pereira et al., 2010b; Adrien et al., 2012). Vacas con dietas pobres en proteína compensan en parte el déficit a través de la movilización de sus reservas corporales y la disminución de la eliminación renal de urea, lo que se refleja en pérdidas de peso, CC y disminución de la producción láctea. Por otro lado, el aumento de proteína plasmática durante el posparto está correlacionado positivamente con el consumo de MS que gradualmente se incrementa durante el posparto (Figura 2C). Las concentraciones de insulina y factor insulino-símil I (IGF-I) están disminuídas alrededor del parto (Figura 2F, Meikle et al., 2004; Cavestany et al. 2009a; Pereira et al., 2010b; Adrien et al., 2012; Astessiano et al., 2012), consistente con la reducción de la ingesta y el BEN que caracteriza este período. Estos datos son consistentes con el conocimiento generado respecto la partición de nutrientes del período de transición de la vaca lechera. La hormona del crecimiento (GH) es la hormona homeorrética o teleoforética por excelencia, que promueve el uso de estos nutrientes por la ubre, ya que «apaga» el consumo del combustible esencial (glucosa) por parte de los tejidos periféricos y promueve la movilización de las reservas corporales. Una de las estrategias utilizadas por la vaca lechera de alta producción es desacoplar el eje somatotrófico, es decir, insensibilización hepática a la GH (menores concentraciones del receptor de GH, GHR) que se traduce en menores concentraciones de su mediador, el IGF-I, (Kobayashi et al., 1999). Resultados nacionales (Astessiano et al., 2012) demuestran que la expresión hepática de los transcriptos de GHR-variante 1A e IGF-I disminuyeron del pre al posparto en vacas lecheras sobre pastoreo, pero en menor medida en vacas alimentadas con raciones totalmente mezcladas (TMR). El desacople del eje GH-IGF-I repercute en varios órganos y tejidos, pero se destaca que la falta de retroalimentación negativa sobre la GH, promueve una mayor concentración circulante de esta hormona, y por lo tanto una mayor acción teleoforética. Asimismo, menores concentraciones de IGF-I e insulina (Figura 2F), favorecen el catabolismo periférico que soporta la lactancia.
Por otro lado, insulina e IGF-I son las mismas hormonas que estimulan el desarrollo folicular y el reinicio a la ciclicidad ovárica luego del parto. Hemos demostrado que una mejor CC al parto (>3) se asocia con concentraciones de IGF-I más altas y anestros posparto más cortos (Meikle et al., 2004). La relación insulina/GH/IGF-I y el día del nadir de balance energético influencian el crecimiento folicular y primera ovulación posparto (Beam y Butler, 1999). El reestablecimiento del BEN se refleja en el aumento de insulina e IGF-I, (Figura 2F) y se asocia con el comienzo del aumento de la ingesta y con el fin del desacople del eje somatotrófico.
Las vacas primíparas presentan mayor dificultad para recuperarse del BEN, reflejado esto a través del perfil metabólico y endócrino más desbalanceado y peores índices reproductivos que las multíparas (Meikle et al., 2004, 2005; Cavestany et al. 2009a; Adrien et al., 2012), proceso probablemente agravado por el estrés que implica su primera lactancia. Esto puede ser el resultado de que estos animales están en desarrollo, aunque también bajo condiciones pastoriles el efecto de dominancia por la disponibilidad de comida está presente (Grant y Albright, 2001). Además, se ha observado que la actividad de pastoreo durante el acceso a la pastura es baja y se observan muy bajas tasas de bocado en vacas primíparas recién paridas (Chilibroste et al., 2012b).
La inmunodeficiencia que caracteriza al período de transición se puede observar en la disminución marcada de las globulinas (Figura 2E) y está asociada al marcado incremento de las patologías infecciosas reproductivas, mamarias, podales u otras (Blowey, 2005; de Torres, 2010). A nivel internacional hay acuerdo en que la incidencia de mastitis es más alta durante los primeros 70 días de lactación (Seegers et al., 2003), lo que es consistente con estudios nacionales (de Torres, 2010), en los que se encontró que el recuento celular por cuarto es significativamente más alto en el primer tercio de lactancia que en el segundo (odd ratio: 1.58). En un trabajo reciente, Rama et al. (2012) reportan que vacas positivas a Leucosis –es decir con densidades ópticas por encima de los controles positivos incluidos en los kits– presentaron una disminución en estos títulos que representan entre 40-60% de los niveles preparto iniciales (Figura 2E). Se demostró la existencia de falsos negativos desde el día -20 al +60, indicando que debe evitarse tomar muestras para diagnóstico serológico de enfermedades infecciosas en este período, al menos para Leucosis bovina. Además de las consideraciones respecto al diagnóstico, la inmunosupresión existente en este período podría modificar la carga de posibles patógenos presentes en la madre durante el periparto, es decir, el animal podría perder la capacidad de controlarlos, reactivando la infección y originando como consecuencia un aumento de su carga con las consecuencias obvias en el propio animal y en el rodeo.
Otras patologías se ven exacerbadas por el BEN que sufre la vaca lechera durante el período de transición. Ramos (2012) ha demostrado que para las condiciones productivas de Uruguay, las cojeras tienen mayor posibilidad de presentarse en la etapa de lactancia media (60-120 días posparto, dpp) respecto a la temprana (<60 días) o tardía (>120 dpp). Esto es consistente con estudios que demuestran que la condición corporal de las vacas está positivamente correlacionada con el espesor del cojinete plantar, el cual disminuye desde el primer mes de lactancia y alcanza su nadir a los 120 días posparto. La disminución del cojinete plantar – estructura compuesta fundamentalmente de tejido adiposo que disminuye presiones mecánicas en el pie– se asocia al aumento en la prevalencia de varias patologías podales (Bicalho et al., 2009).
Nutrientes: énfasis en memoria metabólica y manejo nutricional diferenciales en el periparto
Los efectos del manejo nutricional sobre la producción de leche (cantidad y calidad) y los parámetros reproductivos han sido reportados exhaustivamente a nivel mundial. Sin embargo, los resultados son variables y frecuentemente contradictorios. Los diseños difieren en el grado de reservas corporales al parto (memoria metabólica), nivel de energía/proteína y fuente de la dieta, inclusión de nutrientes específicos, magnitud y duración del BEN, además de otros factores experimentales como raza, edad, momentos en los cuales se aplica tratamientos nutricionales respecto al período de transición y a la toma de observaciones. Esto dificulta la comparación de resultados y la obtención de conclusiones prácticas de manejo, y es especialmente relevante en estudios sobre pastoreo controlado, debido a que la mayor parte de la información proviene de sistemas de producción de leche en condiciones de estabulación.
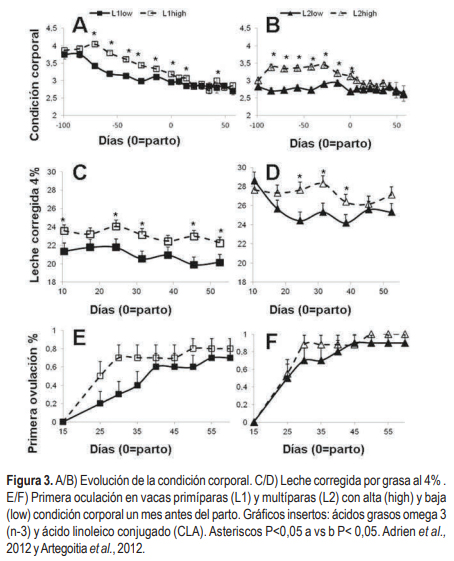
Todos nuestros experimentos han mostrado que vacas con mayor CC movilizan más reservas y presentan mayores concentraciones de NEFA que vacas con pobre CC en el preparto o al parto (Meikle et al., 2004; Adrien et al., 2012). En la Figura 3, se muestra la evolución de las vacas primíparas (L1) y multíparas (L2) con baja (low) o alta (high) CC, inducida nutricionalmente de los -100 a los -30 días preparto (Adrien et al., 2012); a las tres semanas posparto no se encontraron diferencias de CC entre grupos.
Esta mayor oferta de reservas energéticas se visualizó en mayores producciones de leche corregida por grasa y de ácidos grasos polinsaturados (PUFA), como los omega 3 (4,8 vs 2,7 mg/g de ácido graso para alta y baja CC, respectivamente) y CLA (10,3 vs 7,3 mg/g de ácido graso para alta y baja CC, respectivamente) conocidos por su rol beneficioso en la salud humana. Las dietas pastoriles presentan una alta proporción de PUFA (50-75%) de los ácidos grasos totales (Dewhurst et al., 2001), y por lo tanto hemos sugerido que vacas con alta CC presentan un mayor consumo de pasturas que no solo redundaría en contenidos más altos de PUFA ingeridos (Figura 3C, 3D), sino en una tasa de pasaje mayor que limita la hidrogenación ruminal de los mismos (Artegoitia et al., 2012).
Las concentraciones de IGF-I durante el periparto y lactación temprana fueron mayores en las vacas multíparas de CC más alta inducida nutricionalmente (Adrien et al., 2012). Las concentraciones de IGF-I se asociaron a su vez a un reinicio de la ciclicidad posparto más corto; las vacas con alta CC presentaron un anestro 15 días más corto que las de baja CC (Figura 3E, 3F, Adrien et al., 2012). El reinicio a la ciclicidad ovárica afecta drásticamente los indicadores reproductivos de interés económico. En un estudio poblacional con determinaciones de progesterona en leche dos veces por semana durante los primeros 90 días posparto en aproximadamente 900 vacas lecheras se determinó que si un animal no reinicia la ciclicidad ovárica durante los primeros 60 días posparto presenta un alargamiento del intervalo parto primer servicio de 67 días (datos no publicados).
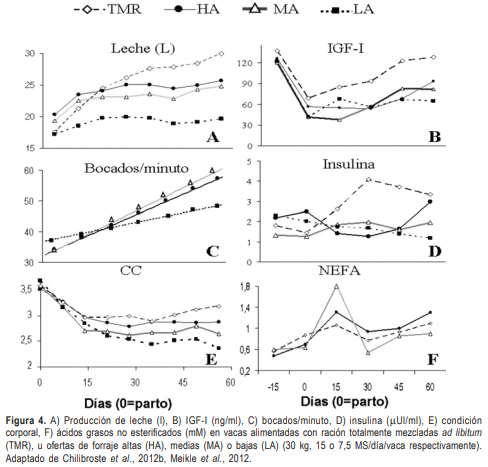
En nuestro país se han realizado numerosos trabajos de suplementación pre y posparto, pero relativamente pocos trabajos incluyeron la determinación del anestro y los perfiles endócrinos. Ofertas de forraje diferenciales (7,5, 15 y 30 kg de materia seca (MS)/vaca/día) y TMR proporcionada ad libitum provocaron niveles de producción de leche y CC acorde a la oferta de nutrientes (Figura 4A, E, Chilibroste et al., 2012b).
En dos experimentos distintos, se observó que la producción de las vacas TMR fue mayor que los grupos a pastoreo y esto fue consistente con una mayor densidad de nutrientes (Meikle et al., 2012; Fajardo et al., 2012), reflejándose en mayores concentraciones de insulina e IGF-I (Figura 4B, 4D; Meikle et al., 2012). No se encontraron diferencias en producción de leche entre vacas con altas (HA) y medias (MA) ofertas de forraje (30 y 15 kg MS/vaca/día, respectivamente), pero estas fueron mayores a vacas con bajas (LA) ofertas de forraje (7,5 kg MS/vaca/día). La probabilidad de pastoreo aumentó linealmente en vacas HA (0,39 min/100 min /día) y MA (0,44 min/100 min/día) a una tasa más alta que vacas LA (0,22 min/100 min/día (Chilibroste et al., 2012b). La tasa de bocado fue mayor en vacas HA y MA respecto LA. La baja de oferta de forraje modificó el comportamiento ingestivo ya que pastorearon menos tiempo y a menores tasas de bocado. Esto se reflejó a partir de la segunda semana posparto en la producción de leche, la cual fue menor que en los otros grupos (Figura 4A). Sin embargo, el grupo de media oferta (MA) mantuvo una producción de leche comparable con el grupo de alta (HA), y observando el perfil endocrino-metabólico (Figura 4B, 4D) se puede sugerir que compensó esta producción con una movilización mayor de reservas (Figura 4E). Las concentraciones más altas de NEFA (Figura 4F) y más bajas de urea (datos no mostrados) en el grupo MA respecto vacas TMR y HA durante el primer mes posparto fue consistente con el reinicio a la ciclicidad ovárica posparto demorado en vacas MA (datos no mostrados).
Otros reportes nacionales han utilizado la suplementación estratégica para la mejora de los índices reproductivos. Cavestany et al. (2009a), suministrando una suplementación energética durante tres semanas preparto a vacas multíparas, reportaron una mayor producción de leche y un anestro 12 días más corto en el grupo suplementado, asociado a una mayor concentración preparto de IGF-I, insulina y leptina. Estos resultados se confirmaron en un experimento posterior (Cavestany et al., 2009b), donde una suplementación energética durante tres semanas preparto redujo la longitud del anestro preparto en vacas multíparas en 15 días, aunque no hubo efecto en vacas primíparas. Por otra parte, la adición de la semilla de girasol a la dieta durante los primeros dos meses posparto promovió la ovulación en vacas primíparas, pero esto no se observó en vacas multíparas (Mendoza et al., 2008), confirmando una vez más la relevancia de la categoría animal en el desempeño reproductivo, aún en respuesta a tratamientos nutricionales. Los autores atribuyeron estos efectos a una densidad energética mayor y/o a la presencia de elementos nutracéuticos en la semilla del girasol.
Líneas genéticas, cruzas y marcadores moleculares
En función de las consideraciones realizadas, respecto de los objetivos de producción definidos en nuestro país, resulta de interés la evaluación comparativa productiva y reproductiva de vacas de diferentes líneas genéticas. Se estudió el desempeño de vacas Holando Uruguayo (HU) y cruza Holstein Friesien Neocelandesa (HU-HFNZ) seleccionadas por mérito económico en la lactancia previa en un mismo ambiente productivo. La producción acumulada de leche, grasa, proteína y sólidos totales a 305 días de lactancia no fueron afectadas por la línea genética; sin embargo, las vacas HU produjeron 0,5±0,23 L/d más de leche que las HU-HFNZ en los primeros 240 días de lactancia (Pereira et al., 2010a). Los porcentajes de grasa y lactosa fueron mayores y los de proteína tendieron a ser mayores en las vacas HU-HFNZ que en las HU. Las vacas HU fueron 49±3 kg más pesadas que las HU-HFNZ y produjeron menos sólidos en leche por kg de peso metabólico. El porcentaje de preñez a los primeros 40 días de servicio fue mayor y la preñez general tendió a ser mayor en las vacas HU-HFNZ que en las HU. Las concentraciones de los ácidos grasos no esterificados y de β-hidroxibutirato aumentaron en torno al parto y fueron mayores en las vacas HU (Pereira et al., 2010b). La eficiencia -producción de sólidos de leche por peso metabólico- fue mayor en las vacas HUHFNZ fundamentalmente debido al menor peso vivo de estas, así como también el desempeño reproductivo. Las diferencias en las variables endócrinas y metabólicas observadas en ambas líneas genéticas sugieren una partición de nutrientes y energía diferente; en la que las vacas HU utilizan mayor energía proveniente de las reservas corporales, mientras que en las HU-HFNZ habría un menor gasto de energía de mantenimiento (Pereira et al., 2010b).
La información generada respecto a cruzas lecheras es incipiente; hemos estudiado el comportamiento productivo y reproductivo de vacas primíparas de hijas de madres Holando Uruguayas y padres Holando americano (HA), y de vacas primíparas cruza Holando Uruguayo con Holando Neozelandés (HNZ), Sueca Roja y Blanca (RBS) y Jersey (J) (Dutour et al., 2010a, 2010b). La producción de leche del grupo HA (17,5±0,37 kg/día) fue superior a las cruzas RBS y J (16,0±0,36 kg/día y 15,1±0,35 kg/día, respectivamente), mientras que no difirió significativamente del grupo HNZ (17,0±0,36 kg/día). Sin embargo, cuando se analizó la producción de leche según el peso metabólico no se encontraron diferencias significativas entre los diferentes grupos genéticos. El grupo J fue significativamente más liviano que el resto de los genotipos, los cuales no variaron entre ellos. No se encontraron diferencias significativas entre los grupos genéticos para varios indicadores reproductivos, pero las cruzas J presentaron mayor proporción de vacas preñadas a los 21 y 42 días de inicio del período de inseminación.
Finalmente y como se mencionó anteriormente, si bien los kits comerciales de marcadores moleculares están disponibles en Uruguay y son utilizados en algunos sistemas de producción, no hemos encontrado estudios al respecto. Nuestros avances respecto a marcadores moleculares, caracteres fenotípicos de interés y endocrinología metabólica se han centrado en la hormona del crecimiento (GHAluI) y su mediador, el IGF-I (IGF-I-Snab I) (Ruprechter et al., 2011). Estos marcadores no presentaron efectos relevantes en parámetros productivos. Sin embargo, un sitio polimórfico (transición T (alelo A) por C (alelo B) ubicado en la región promotora del gen IGF-I y detectado por la Snab I) se asoció con el intervalo parto-primer servicio en vacas primíparas, presentando las vacas genotipo BB intervalos más largos (Ruprechter et al., 2011). Estos resultados son consistentes con estudios anteriores en los que las vacas BB presentaron anestros más largos (datos no publicados). Los hallazgos más relevantes se centraron en los efectos encontrados de los genotipos de GH e IGF-I sobre los perfiles endócrinos y metabólicos, constituyendo el primer reporte internacional que encuentra asociación en vacas lecheras durante el período de transición.
Consideraciones finales
De este documento se desprende la complejidad del sistema biológico que implica la vaca lechera en transición y el medio ambiente productivo. Se demuestra que el manejo nutricional y la genética tienen impactos de relevancia en la endocrinología metabólica durante el periodo de transición que se asocia con desempeños reproductivos diferenciales. Se debe considerar que contribuir en la mejora de un parámetro productivo no implica necesariamente la mejora de la eficiencia económica. Aspectos que usualmente no se toman en cuenta, tales como las variables reproductivas, la salud animal, así como también el mantenimiento a largo plazo del ecosistema pastoril, son parte de la respuesta existente a los manejos realizados por el hombre en el sistema lechero.