Introducción
La grasa en leche de vaca se modifica con facilidad, su concentración depende en particular de la alimentación, y disminuye con niveles bajos de FDN o niveles superiores a 5 % de ácidos grasos (AG) poliinsaturados en la dieta. Estas condiciones favorecen la reducción de grasa en leche debido a dos cambios en la fermentación ruminal. El primero, existe menor síntesis de ácido acético, precursor de síntesis de grasa en la glándula mamaria. El segundo, favorece la síntesis del isómero trans-10, cis-12 del ácido linoleico conjugado (ALC) que inhibe la expresión de enzimas involucradas en la síntesis de AG (Bauman et al., 2011). Dicho isómero es considerado el principal responsable de la reducción de grasa en leche de vacas, y esto se conoce como síndrome de depresión de grasa láctea (SDGL), caracterizado por una reducción hasta 50 % en la concentración de grasa en leche (Bauman et al., 2011). El SDGL se presenta tanto en sistemas de producción estabulados como de pastoreo, aunque con frecuencia mayor en el primero. La desventaja del SDGL se basa en un menor rendimiento de subproductos como queso y mantequilla (Schiavon et al., 2016), y para el productor representa menor precio por la leche. Ante este escenario, se propusieron alternativas para aliviar el SDGL en vacas estabuladas (Vyas et al., 2013) y en pastoreo (Ramírez-Mella et al., 2013) con resultados no satisfactorios. Entonces, se requieren alternativas nutricionales nuevas que contribuyan a la disminución del SDGL. Al respecto, el ácido palmítico (AP) es una opción viable porque incrementa la producción de leche y la concentración y rendimiento de grasa láctea (Piantoni et al., 2013). Con base en este antecedente, la hipótesis en el estudio fue que la adición de fuentes purificadas de AP en dietas para vacas en pastoreo, donde se incluye el isómero trans-10, cis-12 ALC, podría disminuir el SDGL, sin afectar el comportamiento productivo, objetivo principal de la presente investigación.
Materiales y métodos

El experimento se realizó en un rancho ubicado en el municipio de Huimanguillo, Tabasco. El clima se clasifica como cálido húmedo, lluvioso en verano y otoño. La lluvia promedio anual es 2 295 mm año-1, húmedo de junio a octubre con 70 % de la lluvia anual, y seco de noviembre a mayo; la temperatura máxima, media y mínima registrada 35, 25 y 15 °C, respectivamente, con 77.4 % de humedad relativa.
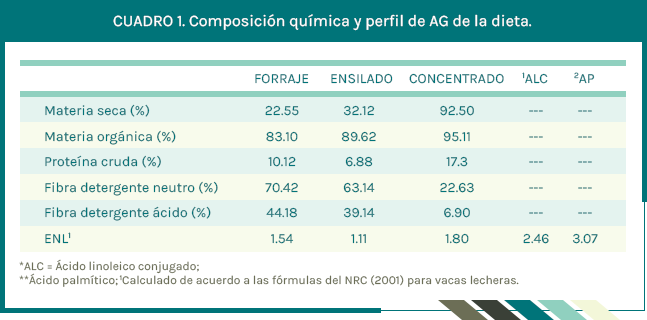
Se utilizaron seis vacas ¾ Pardo Suizo Americano X Cebú en inicios de lactancia (31 ± 7 d en leche y 2.3 ± 0.8 partos; media ± desviación estándar), distribuidas en 3 grupos de 2 vacas cada uno. Los tratamientos se asignaron aleatoriamente a cada uno de los tres grupos, en un Diseño Cruzado. Las vacas tuvieron un periodo de adaptación de 14 d y tres periodos experimentales de 21 d. Los tratamientos fueron: 1) Testigo (Dieta base = pastoreo + alimento concentrado + ensilado de sorgo; Cuadro 1); 2) ALC (dieta base + 50 g de ALC; 3) ALC + AP (dieta base + 50 g de ALC + 412 g de AP). El ALC fue una mezcla de AG microencapsulados, que aportaron 6 g de cis-9, trans-11 y 6 g de trans-10, cis-12 ALC y el AP fue una mezcla de AG microencapsulados > 85 % en AP.
Las vacas estuvieron en pastoreo rotacional intensivo. La pradera fue mixta de Cynodon plectostachyus (18 %), Brachiariadecumbens (12 %) y Brachiaria brizantha (70 %). El consumo de forraje se calculó utilizando óxido de cromo (Cr2O3) como marcador externo y cenizas insolubles en ácido como marcador interno (Geerken et al., 1987). Cada vaca recibió 4 kg d-1de concentrado (mitad en cada ordeño; 05:00 y 16:00 h) y se mezclaba con el ALC y AP para el tratamiento correspondiente. Después de cada ordeña cada vaca recibió 4 kg d-1 de ensilado de sorgo.

Remates Feria

La producción de leche se midió en cada vaca en los últimos 4 d de cada periodo experimental. De cada vaca en cada ordeña se obtuvieron muestras de leche que se almacenaron en viales de plástico (120 ml) y se conservaron en refrigeración a 5 °C para después analizar su composición química.
La determinación de grasa, proteína y lactosa en leche se realizó por espectrometría infrarroja (LactiCheck, EE.UU). Los análisis de las muestras de alimento fueron: MS, PC, cenizas, FDN, FDA (Van Soest et al., 1991), y cenizas insolubles en ácido (Van Keulen y Young, 1977). En las muestras de heces se determinó cenizas insolubles en ácido y concentración de cromo.
La extracción de AG de la leche se realizó según la metodología de Feng et al. (2004). El perfil de AG de los componentes de la dieta y la leche se determinó utilizando la técnica de metilación de Palmquist y Jenkins (2003), en el cual los AG se presentan en forma de metil ésteres. Los metil ésteres de AG se identificaron en un cromatógrafo Hewlett Packard 6890 con inyector automático con una columna capilar de sílice (100 m x 0.25 mm x 0.20 μm de grosor, Sp-2560, Supelco). La identificación de los AG se realizó comparando los tiempos retención de cada pico obtenido del cromatograma, con un estándar de 37 componentes de metil ésteres de AG, y un estándar específico para isómeros cis-9, trans-11 y trans-10, cis-12 de la compañía Nu-Check.
El consumo de energía neta (ENI; MJ d-1) se calculó multiplicando el consumo de MS por la densidad energética de los ingredientes de la dieta (NRC, 2001). La EN para mantenimiento (ENM; MJ d-1) se calculó como el peso metabólico (PV0.75) × 0.08 × 4.184 (NRC, 2001). La EN requerida para la lactancia (NEL; MJ d-1) se calculó como la producción de leche (kg) x [(0.00929 × grasa g kg-1) + (0.00563 × proteína g kg-1) + (0.00395 × lactosa g kg-1)] x 4.184 (NRC, 2001). Dado que las vacas estaban en pastoreo, se consideró el gasto energético por actividad física (ENW; MJ d-1), calculado como la distancia de recorrido diario [(km × 0.0019 × PV) + (0.0050 × PV)] (NRC, 2001). El balance de energía se calculó como ENI – (ENM + ENL + ENW) y se expresa en MJ d-1.
El diseño experimental fue un Diseño Cruzado, y los resultados se analizaron utilizando el procedimiento MIXED de SAS. Las medias se compararon con la prueba de Tukey, se declararon diferencias cuando P ≤ 0.05.
Resultados
El consumo de forraje y MS total no fueron diferentes (P > 0.05) entre tratamientos, pero la producción de leche aumentó (P ≤ 0.05) 1.17 y 1.07 kg d-1 al proporcionar a las vacas ALC + AP respecto al tratamiento testigo y ALC (Cuadro 2).
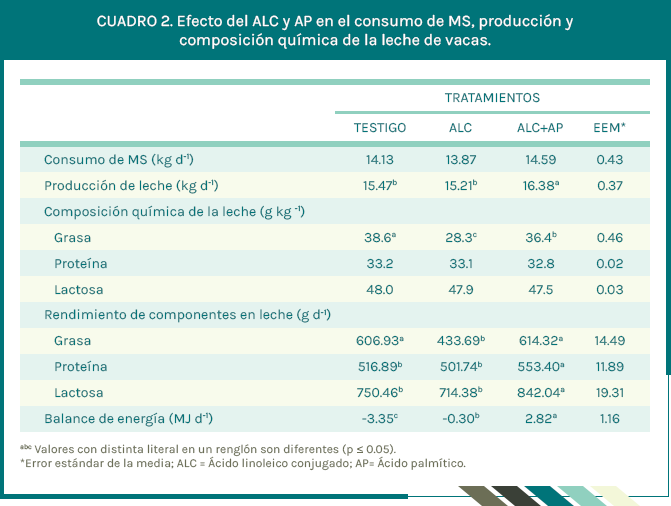
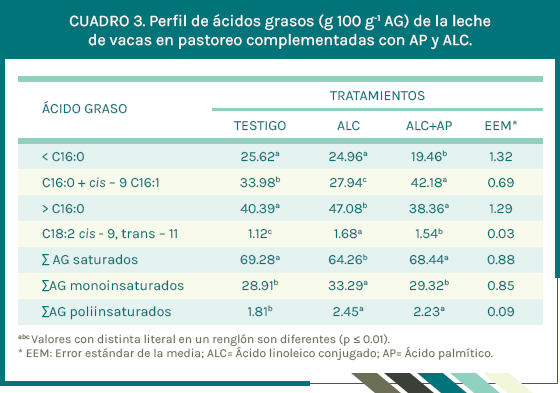
Hubo diferencias (P ≤ 0.05) en el rendimiento de proteína y lactosa en leche (Cuadro 2). La concentración de grasa en leche con los tratamientos ALC + AP y ALC disminuyeron 5.7 % y 26.7 % respecto al tratamiento testigo. Esta diferencia implica que la inclusión de AP en la dieta para vacas en pastoreo disminuyó en 76.7 % la caída de grasa en leche causado por la adición de ALC en la dieta. El balance energético fue diferente (P ≤ 0.05) entre tratamientos, la adición de ALC + AP causó un balance positivo (2.82 MJ d-1) en las vacas, comparadas con el balance negativo de las vacas en los tratamientos testigo (-3.35 MJ d-1) y ALC (-0.30 MJ d-1). La concentración de AG de cadena corta y media (C4:0 a C14:0) en la leche de vacas complementadas con ALC + AP fueron menores (P ≤ 0.05) respecto a los tratamientos testigo y ALC (Cuadro 3).
Discusión
El consumo de MS (Cuadro 2) de las vacas en estudio, está dentro del rango propuesto por el NRC (2001), para vacas con producción de leche inferior a 20 kg d-1, lo cual explica el hecho que no se encontraran diferencias, debido a que la producción máxima de leche en el presente estudio fue 16.38 kg d-1, por lo cual no se esperaría un consumo de MS superior a 16 kg d-1 (NRC, 2001).
La producción de leche aumentó en vacas que recibieron AP. Este resultado es similar al reportado por Piantoni et al. (2013) quienes proporcionaron 384 g de AP a vacas. Sin embargo, Lock et al. (2013) y Rico et al. (2014) no observaron cambios en la producción de leche por efecto del AP, a pesar de haber usado cantidades de AP similares. En el presente estudio, el incremento en la producción de leche con los tratamientos AP se debió al mayor aporte de energía en la dieta por la adición de AP (Cuadro 1). Asimismo, el aumento en la producción de leche de las vacas complementadas con ALC + AP, se debió a un uso más eficiente de glucosa (Hötger et al., 2013). Al respecto, la reducción de síntesis de novo de AG en glándula mamaria provocado por AG de cadena larga, disminuye el uso oxidativo de glucosa en la ruta metabólica de la pentosa fosfato, al inhibir la acetil CoA carboxilasa (Palmquist y Jenkins, 1980), y dado que la glucosa es necesaria para la síntesis de grasa láctea, ya que provee el NADPH empleado en la síntesis de novo, al reducirse la síntesis de novo de AG, existe ahorro de glucosa, misma que puede ser usada para sintetizar más lactosa (Hötger et al., 2013), y considerando que la lactosa es el principal osmo-regulator en la captación mamaria de agua (Rigout et al., 2002), entonces es de esperarse que la producción de leche aumente, como observamos en el presente estudio. El hecho que las vacas de dichos tratamientos tuvieran un ahorro de glucosa, conduce a un uso más eficiente de la EM, lo cual deriva en un balance de energía positivo (von Soosten et al., 2012).
En el tratamiento con solo ALC, la grasa fue menor (P ≤ 0.05) respecto a los tratamientos testigo y AP. Esto indica trasferencia positiva del isómero trans-10, cis-12 ALC de la dieta a leche. El isómero trans-10, cis-12 ALC no afecta la cantidad de grasa en leche en las primeras etapas de la lactancia (Pappritz et al., 2011), debido a que, durante este período, los AG presentes en la leche provienen mayormente de la movilización de las reservas corporales de la vaca, más que de la síntesis de novo, y los efectos del trans-10, cis-12 ALC son más importantes en la síntesis de novo que en la movilización y transporte de AG (Baumgard et al., 2002). Por tanto, el mayor efecto a la síntesis de novo indica el porqué de la eficiencia del AP en la reducción de la caída de grasa por efecto de incluir ALC en la dieta. Esto se debe a que incluir suplementos de AP en la dieta para vacas, como en este estudio, incrementan la disponibilidad de AP en la glándula mamaria. En consecuencia, se reduce la síntesis de novo debido a que la síntesis de grasa láctea a partir de AG dietarios resulta en una ruta metabólica de mayor eficiencia (Loften et al., 2014). Asimismo, el AP estimula la síntesis de triglicéridos en glándula mamaria de vacas (Hansen y Knudsen, 1987). Debido a lo anterior, la concentración de grasa en leche se incrementa (Piantoni et al., 2013; Lock et al., 2013; Rico et al., 2014).
Incluir en la dieta para vacas lactantes AP es más eficiente en estimular la síntesis de AG durante el SDGL provocado por el isómero trans-10, cis-12 ALC, debido a que la trasferencia del AP a leche (Vyas et al., 2013) es hasta del 50 %. Al respecto, Kadegowda et al. (2008) y Vyas et al. (2013) reportan que suplementar con AG no mitiga el SDGL provocado por el isómero trans-10, cis-12 ALC, nuestros resultados son diferentes a los de ellos debió a que la concentración de AP (86.0 %) que utilizamos fue mayor respecto a la utilizada por ellos (27.5 %), lo que llevó a que en nuestro estudio si se mitigara el SDGL. Asimismo, la inclusión de AP en la dieta de las vacas aumentó 29 % la concentración de dicho AG en grasa láctea respecto al tratamiento testigo, indicando trasferencia positiva del AP complementado, lo que en consecuencia provocó incremento en la concentración de grasa en leche.
En el presente estudio, la concentración de ácido palmítico en leche de vacas complementadas con AP fue de 39.5 % similar al reportado por Piantoni et al., 2013; Lock et al., 2013 y Rico et al. (2014), quienes suplementaron AP a vacas lecheras estabuladas. Es interesante observar que a pesar que la dosis de AP en dietas para vacas fluctúa de 500 g a 1,500 g, la concentración de AP en la grasa láctea no rebasa el 40.0 %, aspecto importante a considerar, ya que un valor superior afectaría negativamente la fluidez de la leche dentro de la glándula mamaria (Loften et al., 2014). Sin embargo, esta necesidad de mantener la fluidez en leche, trae como consecuencia disminución de la concentración del isómero cis-9, trans-11 ALC en la leche (Rico et al., 2014), razón por la cual, en el presente estudio, la concentración de dicho isómero en la leche de vacas del tratamiento ALC + AP, fue 8.3 % menor respecto al tratamiento ALC. Sin embargo, al comparar la concentración del isómero cis-9, trans-11 ALC con el tratamiento testigo, fue superior en 72.7 %. Esto indica que la adición de AP a la dieta de vacas complementadas con ALC, incrementa la concentración de ambos AG en la leche, respecto al tratamiento testigo, permitiendo ofrecer al consumidor una leche con un perfil lipídico con mayor potencial funcional.
Conclusión
La inclusión de ácido palmítico en la dieta de vacas ¾ Suizo Americano X Cebú en pastoreo mitiga la caída de grasa en leche provocado por la inclusión de ácido linoleico conjugado en la dieta de las vacas. Además, aumenta el rendimiento de proteína y lactosa en leche, y propicia un balance de energía positivo, pero no cambia el consumo de materia seca ni la concentración de proteína ni lactosa en leche.